Беседы о третьем элементе
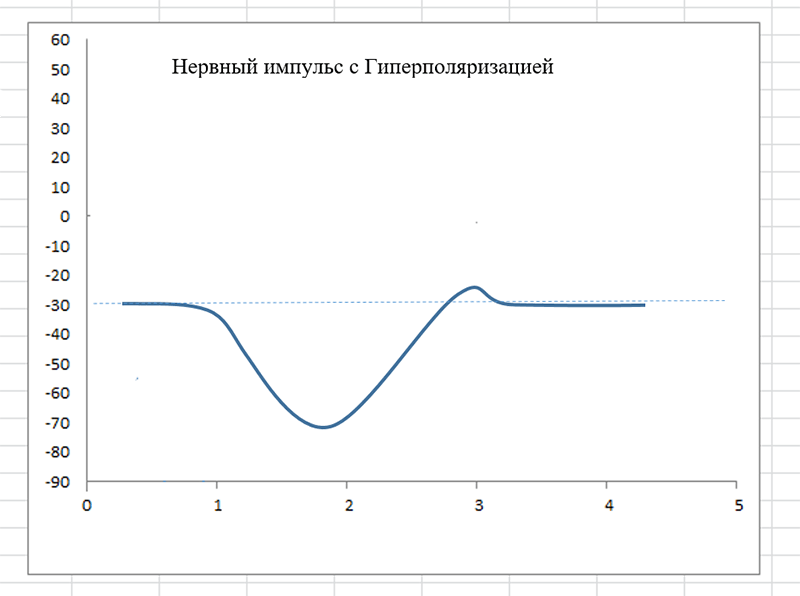
В палочковых и колбочковых нейронах процесс нетипичен (я об этом говорил выше). В них потенциал покоя внутри клетки около -30 милливольт, а при возбуждении он не снижается, а нарастает до -70. Это — так называемая
Осталось рассказать про
Синапсы бывают
Под действием света молекулы родопсина и йодопсина, содержащиеся в рецепторных клетках (палочках и колбочках) меняют свою структуру (геометрию расположения атомов), что приводят в действие многоступенчатую систему химических реакций, запускающих процессы изменение электрического потенциала на мембране этих клеток, к которой подключены дендриты нервных клеток, следующего, биполярного слоя.
В месте соединения имеется типичный
Углубляться в подробный разбор синапсов не стоит, потому что у них великое множество подвидов, и, если все это разбирать досконально, выйдет пять толстых томов. Единственное, что хотелось бы добавить, — запоминание информации и обучение нейронных сетей происходит именно на синапсах. Его свойства обуславливает реакцию клетки адресата на сигнал, пришедший от клетки отправителя, и характер создаваемого синапса приводит к обучению системы.
Несмотря на бесконечную точность, аналоговый сигнал крайне неудобен для передачи сообщений. Во-первых, любой сигнал затухает и расплывается с расстоянием, а также подвержен влиянию различных физических факторов. Бесконечная точность
Ощутив аналоговый сигнал, пришедший снаружи, рецепторный нейрон не может просто отослать его, как есть, в мозг; с сигналом надо что-то сделать, чтобы сообщение не потерялось в шуме и дошло в целости и сохранности до получателя. Чтобы справиться с проблемой шума и ослабления сигнала с расстоянием, нервные клетки работают по принципу
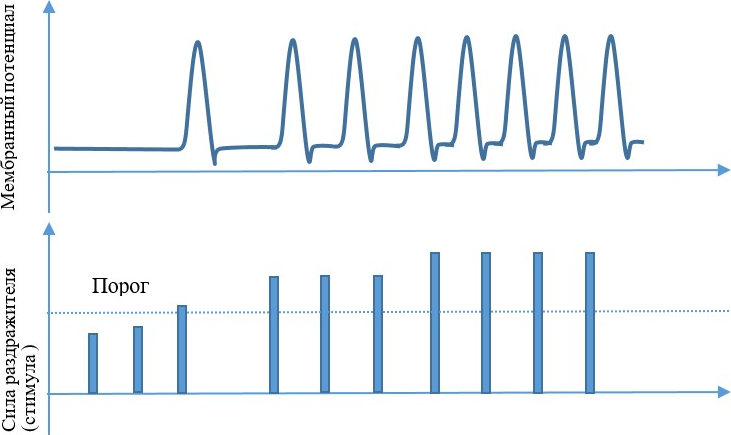
Единичный нервный импульс не несет информации о силе стимула, но зато он как раз такой величины, чтобы его можно было надежно выделить из шума, и не настолько велик, чтобы нас било электрическим током. Но несколько импульсов, пришедших один за другим, вполне могут нам рассказать о силе первичного стимула. Есть некоторый фон по 7–8 импульсов в секунду, означающий, что нейрон на той стороне в принципе жив. Если сигналы идут редко, по 15–20 в секунду, значит стимул был слабый. Но если импульсы барабанят по 150–200 в секунду, значит, стимул сильный, и глаз надо прикрыть, чтобы не ослепнуть.
Матчасть номер два (физиология зрительных путей)
Зрительные нервы обоих глаз перекрещиваются в хиазме и входят в
В сетчатке имеется около миллиона ганглиарных клеток, каждая из которых выпускает один аксон, соответственно, зрительный нерв содержит около миллиона проводков. Получается, миллион пикселей или мегапиксель — это среднее разрешение матрицы нашей сетчатки, но на деле острота зрения гораздо выше, потому что расположен этот мегапиксель очень неоднородно.
В области центральной ямки (желтого пятна) количество рецепторов на несколько порядков выше, чем на периферии, упакованы они крайне плотно и буквально жмутся друг к дружке. Эти нейроны отличаются малыми размерами и называются
В месте выхода зрительного нерва из сетчатки рецепторов нет, и там находится так называемое слепое пятно. Его мы не различаем и если какой-нибудь предмет окажется в его поле, он станет невидим.
Перед входом в первые центральные мозговые структуры зрительные пути проходят стадию перекреста, называемую
Перекрест хиазмы практически полный у тех животных, у которых глаза расположены по бокам головы. Но у обладающих бинокулярным зрением, то есть тех, у кого глаза сдвинуты вперед, так что поля зрения обоих частично или полностью перекрываются, логика перекреста изменена.
Очень удобно обозревать окрестность, когда глаза находятся по бокам от головы, — видно все, что делается спереди, сбоку и сзади. Можно спокойно греться на солнышке и чистить перья. А если какой-нибудь умник попробует незаметно подкрасться сзади, следует нагло зыркнуть черным глазом, злобно каркнуть, показать язык и улететь.
Но если ты прыгаешь с ветки на ветку, ситуация в корне меняется. Главной проблемой становится уже не обзор окрестностей, а то, что, если не допрыгнешь до следующей ветки на несколько сантиметров, упадешь с дерева и переломаешь себе все кости. В такой ситуации гораздо важнее уметь точно определять расстояние, и тут становится крайне выгодно иметь бинокулярное зрение.
Глаза сдвигаются на переднюю часть головы, и область обзора перекрывается так, что каждый глаз видит предмет под немного другим углом. Это позволяет сравнивать картинки. Для этого в хиазме половина нервных волокон уже не перекрещиваются, а остаются на своей стороне, причем волокна от одинаковой стороны обоих глаз оказываются в одном пучке, вместе. Сравнивая ту же самую картинку, пришедшую под разными углами зрения, мозг находит расстояния до объектов, достраивая воображаемый треугольник.